
Термин
«мутация»впервые был предложенГ.
Де Фризомв его классическом труде
«Мутационная теория» (1901-1903).
Основные положения мутационной теории:
1.Мутация
возникает скачкообразно, т.е.
внезапно, без переходов.
2.
Образовавшиеся новые формы наследуются,
т.е. являются стойкими.
3.
Мутации не направлены(т. е. могут
быть полезными, вредными или нейтральными).
4.
Мутации – редкие события.
5. Одни
и те же мутации могут возникать повторно.
Мутация
– это скачкообразное стойкое
ненаправленное изменение генетического
материала.
3.
Закон гомологических рядов в
наследственной изменчивости
Следующим
после мутационной теории Де Фриза
серьезным исследованием мутаций была
работа Н.И. Вавилова по наследственной
изменчивости у растений.
Изучая
морфологию различных растений, Н.И
Вавиловв1920 г. пришел к выводу,
что, несмотря на резко выраженноеразнообразие(полиморфизм) многих
видов, можно заметить и четкиезакономерностив их изменчивости.
Если взять для примера семейство злаков,
то окажется, что одинаковые отклонения
признаков присущи всем видам (карликовость
у пшеницы, ржи, кукурузы; колоски
безостые, неосыпающиеся и т.д.).
Закон
Н. И. Вавиловагласит: «Виды и роды,генетически близкие, характеризуютсясходными рядами наследственной
изменчивостис такой правильностью,
что, зная ряд форм в пределах одного
вида, можнопредвидеть нахождение
параллельных форму других видов и
родов».
Свой
закон Н.И. Вавилов выразил формулой:
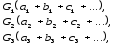
где
G1,
G2,
G3,– виды, аa, b,
c– различные
варьирующие признаки.
Этот
закон важен прежде всего для селекционной
практики, потому что даёт направление
поиска неизвестных форм у растений
(вообще у организмов) данного вида, если
они уже известны у других видов.
Под
руководством Н.И.Вавилова были организованы
многочисленные экспедиции по всему
миру. Из разных стран были привезены
сотни тысяч образцов семян культурных
и диких растений для коллекции Всесоюзного
института растениеводства (ВИР). Она до
сих пор является важнейшим источником
исходных материалов при создании новых
сортов.
Теоретическое
значениеэтого законасейчасне кажется столь большим, каким считалось
в 1920 г. В законе Н.И. Вавилова содержалось
предвидение того, что у близкородственных
видов должны бытьгомологичные,
т.е. сходные по структуре гены. В тот
период, когда о структуре гена ничего
не было известно, это был, безусловно,
шаг вперед в познании живого (закон
Н.И.Вавилова сравнивали по значимости
с периодическим законом Д. И. Менделеева).
Молекулярная генетика, секвенирование
генов подтвердили правильность догадки
Н.И. Вавилова, его идея стала очевидным
фактом и уже не является ключом к познанию
живого.
4. Классификация мутаций
Наиболее
полную классификацию мутаций предложил
в 1989 г.
С. Г. Инге-Вечтомов. Приводим
ее с некоторыми изменениями и дополнениями.
I.
По характеру изменения генотипа:
-
Генные
мутации, или точковые. -
Хромосомные
перестройки. -
Геномные
мутации.
II.
По характеру изменения фенотипа:
-
Морфологические.
-
Физиологические.
-
Биохимические.
-
Поведенческие
III.
По проявлению в гетерозиготе:
-
Доминантные.
-
Рецессивные.
IV.
По условиям возникновения:
-
Спонтанные.
-
Индуцированные.
V.
По локализации в клетке:
1.
Ядерные.
2.
Цитоплазматические (мутации внеядерных
генов).
VI.
По возможности наследования (по
локализации в организме):
1.
Генеративные (возникшие в половых
клетках).
2.
Соматические (возникшие в соматических
клетках).
VII.
По адаптивному значению:
-
Полезные.
-
Нейтральные.
-
Вредные
(летальные и полулетальные).
8.
Прямыеиобратные.
Теперь
дадим пояснения по некоторым типам
мутаций.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Описание презентации по отдельным слайдам:
-

1 слайд
Министерство здравоохранения
Петровск – Забайкальский филиал
ГПОУ «Читинский медицинский колледж»МУТАЦИОННАЯ ТЕОРИЯ
ГУГО ДЕ ФРИЗА
Подготовила: преподаватель
Николаева Ольга Владимировна2023 г
-
2 слайд
ВИКИПЕДИЯ
Гуго Де Фриз. Голландский ботаник. Один из основателей генетики. Родился в 1848 году, умер в 1935 году.
Получил образование в Лейденском университете, с 1866 года изучал
там ботанику и защитив в 1870 году дипломную работу о влиянии тепла
на корни растений, несколько месяцев слушал лекции по химии и физике
в Гейдельбергском университете и работал в лаборатории Юлиуса Закса
в Вюрцбурге.
В отличие от многих других ученых, он не оставил после себя научной
школы. Но его работы составляют важный этап в истории науки.
Основные положения теории мутаций изложил Г.де Фриз в работе
«Мутационная теория»(1901-1903). Ввел термин мутация.
https://ru.wikipedia.org/wiki/Де_Фриз,_Хуго -
3 слайд
ОПРЕДЕЛЕНИЕ
Мутации – стойкое (которое может быть унаследовано потомками данной клетки или организма) изменение генома.
Термин предложен Гуго де Фризом в 1901 году.
Это естественное природное явление, закрепленное отбором.
Свойство живых организмов обусловлено структурой генетического
материала и многоэтапностью основных биологических процессов,
в которых принимают участие факторы наследственности.
Мутации изменяют количество и структуру ДНК, которые
определяются измененными генами.
https://yandex.ru/images/search?rpt -
4 слайд
ОСНОВНЫЕ ПОЛОЖЕНИЯ МУТАЦИОННОЙ ТЕОРИИ
1.Мутация возникает скачкообразно, то есть внезапно, без переходов.2. Образовавшиеся новые формы наследуются, то есть являются стойкими.
3. Мутации не направлены(то есть могут быть полезными, вредными или нейтральными).
4. Мутации – редкие события.
5. Одни и те же мутации могут возникать повторно.
-
5 слайд
ОШИБКА МУТАЦИОННОЙ ТЕОРИИ ДЕ ФРИЗА
Он противопоставил теорию мутаций, теории естественного отбора.
Де-Фриз не правильно считал, что мутации могут сразу дать образование новых видов.
По де-Фризу прогресс эволюции связан с коротким
промежутком времени то есть мутационным
периодом.
https://topslide.ru/ -
6 слайд
ИНФОРМАЦИОННЫЕ ИСТОЧНИКИ
Рубан Э.Д. Учебник «Генетика человека с основами медицинской этики», 2016
Интернет источники:
— ru.wikipedia,.org;
— biografiivsem.ru;
— studfile.net;
— lektsii.org.;
— https://topslide.ru/;
— https://Yandex.ru/images;
— https://ru.wikipedia.org/wiki/Де_Фриз,_Хуго/ -
7 слайд
СПАСИБО ЗА ВНИМАНИЕ







Биология
Тема 4: Видеоуроки. Основы генетики
Урок 15: Мутационная теория. Причины мутаций
- Видео
- Тренажер
- Теория
Заметили ошибку?
Эволюционное значение мутаций
Мутация может иметь эволюционное значение только при условии передачи ее по наследству. Для этого мутация должна произойти в предшественниках половых клеток. Сами половые клетки: для животных это яйцеклетка и сперматозоид будут носителями такой мутации (рис. 1).

Рис. 1. Предшественники половых клеток и половые клетки
Мутации в половых клетках называются генеративными, мутации в прочих клетках называются соматическими.
Соматические мутации не играют большой роли в эволюции, так как они не наследуются. Причины, приводящие к возникновению мутаций, вызываются факторами, которые повреждают молекулы ДНК (рис. 2), это ультрафиолет, радиационное излучение и высокие температуры.

Рис. 2. Факторы, повреждающие молекулы ДНК
Кроме того, мутации вызываются специальными химическими веществами, которые называются мутагены (рис. 3).

Рис. 3. Мутаген
Мутагены могут поступать в организм с пищей или вдыхаемым воздухом, они также могут образовываться и внутри организма. Наиболее типичный внутренний мутаген – это перекиси водорода, образующиеся при дыхании.
Мутации возникают не только при повреждении самой ДНК, но и при повреждении механизмов копирования и восстановления ДНК. Частота мутаций, обусловленных внутренними причинами, более или менее постоянна, она составляет около одной мутации на копирование десяти миллионов пар нуклеотидов – у взрослого мужчины около 10–15 % сперматозоидов (рис. 4) имеют как минимум одну новую мутацию.

Рис. 4. Сперматозоиды
Разные регионы генома имеют разную частоту возникновения мутаций, в высоко мутабельных регионах частота гораздо выше, чем в консервативных.
В природе существуют механизмы, которые специально увеличивают скорость мутаций, например в иммунных клетках (рис. 5).

Рис. 5. Лимфоцит
Для успешного иммунного ответа требуется наличие очень большого разнообразия антител, чтобы при каждой новой инфекции можно было быстро подобрать нужное антитело. Для этого клетки, синтезирующие антитела, должны часто мутировать, чтобы антитела постоянно отличались друг от друга. Во многом эффективность нашего иммунитета зависит от скорости мутаций иммунных клеток. А частота мутаций, вызванных внешними факторами, зависит в первую очередь от интенсивности самого фактора и индивидуальной устойчивости.
Мутационная теория Хуго де Фриза
Представление о мутациях достаточно полно сформулировал голландский ботаник Хуго де Фриз (рис. 6).

Рис. 6. Хуго де Фриз
Эти положения сейчас называются мутационной теорий де Фриза, она состоит из следующих тезисов:
- Мутация – это дискретное изменение наследственного материала.
- Мутация – это редкое событие. Вероятность обнаружения мутаций зависит от числа исследованных особей.
- Мутации устойчиво передаются из поколения в поколение.
- Мутации возникают ненаправленно (спонтанно) и, в отличие от модификаций, не образуют непрерывных рядов изменчивости.
Мутации создают основу для естественного отбора, в результате мутаций появляются особи с резко отличающимися признаками (рис. 7).

Рис. 7. Альбинос
Как правило, такие признаки неудобны и приводят к элиминированию особей, но иногда эти признаки оказываются полезными и такая особь дает начало новому виду. Чаще всего полезные мутации возникают при изменчивых факторах внешней среды.
Список литературы
- Мамонтов С.Г., Захаров В.Б., Агафонова И.Б., Сонин Н.И. Биология 11 класс. Общая биология. Профильный уровень. – 5-е издание, стереотипное. – Дрофа, 2010.
- Беляев Д.К. Общая биология. Базовый уровень. – 11 издание, стереотипное. – М.: Просвещение, 2012.
- Пасечник В.В., Каменский А.А., Криксунов Е.А. Общая биология, 10-11 класс. – М.: Дрофа, 2005.
- Агафонова И.Б., Захарова Е.Т., Сивоглазов В.И. Биология 10-11 класс. Общая биология. Базовый уровень. – 6-е изд., доп. – Дрофа, 2010.
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
- Интернет-портал «activestudy.info» (Источник)
- Интернет-портал «big-archive.ru» (Источник)
Домашнее задание
- Какие мутации могут иметь эволюционное значение?
- Назовите причины, приводящие к возникновению мутаций.
- Из чего состоит мутационная теория де Фриза?
Заметили ошибку?
Расскажите нам об ошибке, и мы ее исправим.
Ч. Дарвин разделял положение о том, что природа не делает скачков. Здесь заключался принимаемый дарвинистами постулат о непрерывности и о неограниченности изменчивости. Принималось, что различия между разновидностями и видами обязаны возникновению мелких непрерывных изменений. Хотя Ч. Дарвин знал, что иногда наблюдаются резкие отклонения от нормы, он считал, что эти своеобразные «уродцы» очень редки и в силу слабой жизнеспособности не имеют эволюционной судьбы.
Этот вывод Ч. Дарвина и его последователей был поставлен под сомнение исследованиями Г. де Фриза, который по праву считается основателем мутационной теории, основные положения которой остаются верны и сегодня (Лобашев, 1967, с. 287). К этому можно добавить, что к славе Г. де Фриза как переоткрывателя законов Менделя следует добавить два важных пионерских концептуальных представления: 1) о нестабильном состоянии генов и 2) о существовании критических периодов в проявлении наследственных признаков (Фриз, 1932; Vries, 1912).
Подробно рассматривая динамику идей в области учения о макроэволюции, В. И. Назаров (1991) приходит к выводу, что де Фриз высказал три кардинальных положения, составивших ядро современной теории прерывистого равновесия и находящихся сейчас в фокусе острых дискуссий: идею о неравномерности темпа мутирования и эксплозивности видообразования, представление о двух разных путях видообразования — «групповом» и «филогенетическом» (это близко к современным представлениям о кладистическом и филетическом видообразовании), а также идею независимости акта видообразования от естественного отбора, борьбы за существование.
С точки зрения истории и методологии науки интересно, что нередко исследователь специально отбирает тот материал для исследования, который соответствует его интуитивной гипотезе. Мендель специально выбрал горох как жесткий самоопылитель и специально выбрал для исследования четкие единичные признаки, поведение которых можно было с легкостью изучать в ряду поколений. Он писал, что «выбор растительной группы, которая будет служить опытам подобного рода, должен быть сделан с наивозможной осторожностью, если мы не хотим подвергнуть риску самый успех опыта». Напротив, де Фриз подыскивал для работ по анализу наследственной изменчивости вид растений, который в силу каких-то внутренних причин отличался бы сильной изменчивостью. Только после того, как Г. де Фриз просмотрел 100 видов, он остановился на энотере. И его выбор оказался фантастически удачен для задуманных целей.
На одном заброшенном картофельном поле вблизи деревни Гильверзум в 1886 г. он обратил внимание на популяцию энотеры, растения, ввезенного из Америки и одичавшего в Европе. В следующем 1887 г. он нашел на этом же поле двух мутантов (забегая вперед в терминологии) и заложил многолетний опыт по изучению частоты возникновения мутантов.
Энотера оказалась уникальной в смысле своей генетической конституции, а изучение закономерностей ее наследственной изменчивости и видообразования таит в себе еще множество загадок. Работая с энотерой, Г. де Фриз получил от первых высеянных в 1886 г. девяти растений около 53 тыс. потомков в период 1886–1899 гг. и среди них около 800, или 1,5 % форм, имеющих резкие отклонения от исходного типа. Эти отклонения были названы мутациями. Мутации возникали внезапно, непредсказуемо, в разных направлениях. В ряде случаев, отдельные мутации захватывали сразу множество признаков и полностью изменяли габитус растения, причем эта совокупность признаков передавалась как дискретная единица. Такие резко отличающиеся друг от друга формы, возникающие внезапно, де Фриз назвал «элементарными видами». К их появлению отбор никакого отношения не имеет. Что касается вида в дарвиновском понимании, то он, по мнению де Фриза, представляет собой совокупность «элементарных видов», впоследствии названных линнеоном.
Совершенно сходные идеи в работе «Гетерогенезис и эволюция. К теории происхождения видов» развил необычайно талантливый ученый, директор ботанического сада при Петербургском университете, академик С. И. Коржинский (1861–1900), проживший всего 39 лет. Гетерогенезисом был назван процесс внезапного сальтационного появления форм, отличающихся комплексом новых признаков. «Эти признаки обладают большим постоянством и неизменно передаются по наследству из поколения в поколение. Таким образом, сразу возникает новая раса столь же прочная и постоянная, как и те, которые существуют с незапамятных времен» (цит. по: Филипченко, 1977, с. 162).
Что касается происхождения садовых растений, то С. И. Коржинский категорически утверждал, что во всех известных ему случаях новые разновидности возникли путем внезапных отклонений от исходных форм. В своих лекциях по генетике, опубликованных в Мюнхене в 1911 г. и в русском переводе два года спустя, Рихард Гольдшмидт заключает, что «среди ботаников заслуга накопления наибольшего числа фактических данных, имеющих отношение к внезапному возникновению растительных форм, принадлежит русскому ботанику академику С. И. Коржинскому» (Гольдшмидт, 1913, с. 135).
Инициированные де Фризом исследования по генетике, цитологии и систематике в роде Oenothera, привели к открытию фантастических механизмов в организации генетической системы размножения, способах наследственной изменчивости и видообразования. В сводке по цитогенетике (Суонсон, Мерц, Янг, 1969) ситуация с энотерой образно названа «бунтом против эволюции». Здесь оказалось «можно то, что нельзя», а именно: сочетание таких элементов генетической системы, каждый из которых сам по себе вреден, а в комбинации дает вполне удовлетворительный результат. Среди них: 1) вовлечение группы или целого набора хромосом в реципрокные транслокации, что обычно приводит к резкому падению фертильности вследствие хромосомной нестабильности гамет и зигот; 2) накопление летальных мутаций; 3) резкое ограничение рекомбинаций, когда транслокационный комплекс передается как целое; 4) неспособность целого набора хромосом проходить либо через женский, либо мужской гаметогенез.
И несмотря на это, завезенные два-три века назад в Европу из Северной Америки растения рода Oenothera образовали сложные гибридные формы видового ранга, по терминологии В. Гранта (1984) «гетерогамные микровиды». Они распространились в Европе как сорняки, а вид Ое. lamarckiana, который специально был выбран де Фризом для исследования, не только распространился по всей Европе, а позднее вновь утвердился в Америке!
Распространено мнение, к сожалению кочующее по разным учебникам и сводкам, что де Фриз обнаружил не мутации, а лишь редкие рекомбинации, выщепляющиеся в потомстве транслокационных гибридов. Но это, во-первых, не так. А во-вторых, в случае энотеры трудно отличить мутации и рекомбинации. С современных позиций очевидно, что процессы мутации и рекомбинации на молекулярном уровне переплетены и разграничение это нередко вопрос терминологии (Хесин Р. Б., 1984, с. 279). Г. де Фриз обнаружил три типа мутаций: генные, хромосомные и геномные. Среди типично генных были «brevistytis» с коротким столбиком цветка и nana — карликовые растения (Cleland, 1935,1962; Грант, 1984; Burnham, 1980).
К хромосомным мутантам, обнаруженным Г. де Фризом и его учениками в первые же годы, относятся трисомики (2n = 15 вместо 2n = 14). Сначала де Фриз выделил их как относительно константные формы, отличающиеся множеством признаков, и лишь затем был определен их хромосомный статус как трисомиков.
Он впервые обнаружил формы, которые в терминах классической генетики соответствуют геномным мутациям. А именно, полиплоидную форму gigas, которая возникла на 4-й год его опытов среди около 14000 исследованных растений. И, наконец, в разные годы среди в общей сложности 54 000 исследованных растений де Фриз зафиксировал возникновение так называемых «полумутантов» (частота их появления 0,06 %). Растения этого типа, названные rubrinervis, характеризовались красными жилками на листьях, широкой красной полосой на чашечке и плодах и слабым развитием луба.
Как позднее было показано (Cleland, 1962), кариотип исходного вида Ое. lamarckiana, с которым работал Г. де Фриз, имел 12 пар хромосом, объединенных в транслокационное кольцо, и одну пару свободную. А «полумутант» rubrinervis имел 4 свободных пары и 6 пар, объединенных в кольцо. Для возникновения такой формы в потомстве отдельных растений рекомбинационным путем необходимо, чтобы в отдельных половых клетках одномоментно происходили множественные рекомбинационные обмены, причем в определенных участках! Такое событие можно представить себе только в рамках современной генетики, когда стало известно, что множественные обмены могут происходить в местах локализации мобильных элементов, а также быть следствием так называемых «транспозиционных взрывов» и множественных упорядоченных транспозиций (Engels, 1983; Герасимова, 1990; Гвоздев, Кайданов, 1986, 1990). Можно суверенностью полагать, что подобные события будут описаны и у энотеры, исследование которой сулит множество открытии в области механизмов наследственной изменчивости.
В эволюционной генетике, помимо энотеры, доводы в пользу множественных одномоментных хромосомных перестроек получены при исследовании хромосомного полиморфизма у роющих грызунов — слепушонок группы Ellobius tancrei, относящихся к надвиду Е. talpinus. Здесь были открыты так называемые «робертсоновские веера» с варьированием в числе хромосом от 54 до 32. При этом получены доводы, что «54 и 32-хромосомные кариотипы должны были возникнуть за счет резкой скачкообразной перестройки всего кариотипа в целом, приведшей к одномоментному слиянию 16–20 акроцентрических хромосом и 8–10 метацентриков» (Воронцов, 1988, 1999).
Наконец, с точки зрения путей видообразования де Фриза следует считать первооткрывателем двух новых типов видообразования: 1) хромосомного, в частности полиплоидного и 2) гибридогенного. В случае гибридогенного видообразования один вид содержит два разных хромосомных комплекса, которые передаются потомству как целое и в разных комбинациях скрещивания «порождают» новые виды. Ученик и последователь де Фриза О. Реннер открыл, что геном вида Ое. lamarckiana, с которым работал де Фриз, состоит из двух транслокационных комплексов gaudens и velans. В них все 7 пар хромосом связаны взаимными обменами и передаются совместно как одна гигантская хромосома.
При скрещивании растений вида Ое. lamarckiana (комплексы velans/gaudens) и Ое. strigosa (комплексы deprimens/stringers) получаются все четыре возможные комбинации, имеющие статус вида и распространенные в природных популяциях (Грант В., 1984, с. 368). Другими словами, в каждом скрещивании происходит «порождение видов», которое можно наблюдать воочию!
Открытые у энотеры транслокационные гибридные видовые системы, названные комплексами Реннера, обладают еще двумя поразительными особенностями. Во-первых, летальностью каждого из них в гомозиготе, так что возникает сбалансированная перманентная гетерозиготность, когда выживают только гетерозиготы. Именно этот, открытый в опытах де Фриза феномен, натолкнул Г. Меллера на идею использовать метод сбалансированных леталей для создания генетического метода количественного учета возникновения мутаций у дрозофилы. Во-вторых, один из реннеровских комплексов передается только через яйцеклетку, а другой — только через пыльцевые клетки, т. е. в гаметогенезе у гибридогенного вида происходит элиминация одного из геномов.
Кратко суммируем значение работ де Фриза для генетики видообразования и эволюционной теории. Г. де Фриз установил неравномерность темпа эволюционного процесса, наличие вспышек мутаций, которые происходят в «мутационные периоды». В период этих вспышек возникают скачкообразные мутации, резко меняющие облик растений, вызывающие резкие изменения в системе воспроизведения, в результате чего сальтационно возникают формы видового ранга. Подобные сальтационные изменения могут происходить повторно. В опытах Г. де Фриза и его учеников и последователей на энотере было открыто хромосомное видообразование (полиплоидия) и гибридогенные виды, образованные на основе транслокационных комплексов.
Справедливо считается, что «классическая дарвиновская концепция понимает видообразование как постепенный градуалистический процесс» (Воронцов, 19886). Генетические исследования де Фриза доказали возможность хромосомного сальтационного (полиплоидия) и гибридогенного видообразования. Оно оказалось повсеместно распространенным у растений. У покрытосемянных растений около 52–58 % видов являются полиплоидами, по преимуществу это аллополиплоиды, растения объединяющие геномы разных видов. Причем, с такими сложными переплетениями полового, бесполого, гибридного размножения, с такими поразительными системами естественной хромосомной инженерии, что приходится расширить концепцию вида до разных по емкости видовых единиц (Грант, 1984, Малецкий, 1995).
Ведущее концептуальное открытие в этой области было сделано в 1917 году датским генетиком О. Винге, который предположил, что у межвидовых гибридов в случае удвоения у них числа хромосом, могут восстановиться мейоз и плодовитость. «Настоящее исследование имеет непосредственное отношение к этой идее. Оно может рассматриваться как экспериментальное обоснование теории гибридного происхождения полиплоидных видов», — таково введение к классической работе Г. Д. Карпеченко (1927). Он впервые синтезировал новую неизвестную в природе видовую форму Raphanobrassica, константный полиплоидный межродовой гибрид между редькой и капустой. Совершенно справедливо Н. Н. Воронцов (1999) называет синтез рафанобрассики первым случаем конструирования нового генома, того, что в конце 70-х стало называться генетической инженерией.
Через три года шведский генетик Арне Мюнтцинг впервые осуществил ресинтез дикорастущего в природе аллополиплоидного вида пикульника рода Galeopsis. Природная хромосомная инженерия создает гибридогенные полиплоидные комплексы видов, открытые и изученные американским ботаником Ледьярдом Стеббинсом. В этих комплексах геномы нескольких диплоидных исходных видов могут вступать между собой во всевозможные гибридные аллотетраплоидные комбинации. Объединяться могут сразу несколько геномов, так что предком одного вида может ни один, а несколько видов, как, например, у обычной мягкой пшеницы, у видов хлопчатника (Грант, 1984).
Неожиданно выяснилось, что гибридогенное, сходное в принципе с энотерой, видообразование регулярно встречается в ряде групп позвоночных и беспозвоночных животных. Гибридогенное видообразование описано уже у 50 видов позвоночных (Боркин, Даревский, 1980, 1989; Кирпичников, 1988; Tunner, Heppich-Turmer, 1991; Даревский, 1995). Оно принимает три основные формы: партеногенез (спермин не нужны для развития яйцеклеток видов-гибридов); гиногенез (спермин нужны лишь для активации развития, но развитие происходит на основе женских гамет и наследование матроклинно); и собственно гибридогенез, когда гибридный вид образуется на основе гибридных зигот, но один из родительских геномов селективно элиминируется в клетках полового пути в ходе гаметогенеза и мейоза.
Гибридогенез оказался широко распространен у скальных ящериц рода Lacerta, живущих в горных районов Закавказья. Среди 18 близких видов комплекса L. saxicola 7 имели гибридогенное происхождение. Все они состояли из партеногенетических самок, способных размножаться без самцов. Но они способны также скрещиваться со своими собратьями из двуполых видов и порождать новые гибридогенные виды. В последнем варианте примерно в 10 % случаев возникают триплоиды. Причем у некоторых групп ящериц, вьетнамских гекконов и агам, триплоиды оказались плодовитыми Поскольку гибридогенез здесь является нормой, то (1) одни и те же партеновиды порождаются вновь и вновь в разное время и (2) одни и те же двуполые виды способны вступать в разные гибридогенные связи и порождать таким образом разные виды! (Даревский, Гречко, Куприянова, 1999).
У животных обнаружено также удивительное явление избирательной элиминации одного из геномов получило название «геномное исключение». У гибридного вида хромосомы одного из родительских видов как бы одалживаются для построения соматических тканей. Вот почему было предложено именовать такой способ видообразования как «кредитогенез» (Боркин, Даревский, 1980). Элиминация одного из родительских геномов в клетках полового пути может быть названа также как «геномоцид». Это явление было описано и у растений в случае отдаленных гибридов ячменя. Природный гибридогенез в сочетании с геномоцидом в зародышевом пути был обнаружен у некоторых видов беспозвоночных.
Особенно поразительной оказалась гибридогенная система у палочников рода Bacillus, где сосуществуют пять способов воспроизведения (Giorgi, 1992; Mantovani, Scali, 1992):
1) обычное бисексуальное размножение, 2) гибридогенез, 3) партеногенез, 4) гиногенез и 5) андрогенез — первый случай такого рода, обнаруженный как регулярное событие в природных популяциях (схема на рис. 1).
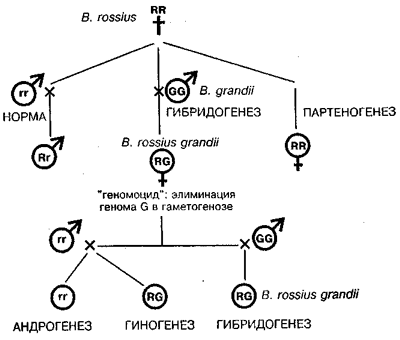
Рис. 1. Гибридогенез у видов палочника (род Bacillus) как пример недарвиновского видообразования, открытого на энотере Г. де Фризом. R и r обозначают геномы вида В. rossius у самок и самцов, G — геном В. grandii (no Mantovani, Scali, 1992; Giorgi, 1992).
В 1900 году Хуго Де Фриз заново открыл законы наследования Грегора Менделя, а спустя год предложил свое объяснение эволюции феноменом мутационизма: один вид переходит в другой скачком, в одну мутацию. Это было верно лишь частично — только для того, что сейчас называют макромутациями, но в начале прошлого века мутационная теория Де Фриза задала вектор для всех дальнейших генетических исследований, так как механизм мутирования определял, что из чего делается в живой природе. Например, сейчас ученым понятно, из чего и как природа производит вирусы, подобные нынешнему коронавирусу, но справиться с кошмаром вирусных мутаций они пока не могут. И это не первый «кошмар» в истории генетики, правда, предыдущий касался чистой теории.
Один монах и три ботаника
В январе 1900 года добрые люди дали почитать голландскому ботанику Хуго Де Фризу статью Грегора Менделя «Versuche uber Pflanzen-Hybriden» («Опыты над гибридами растений») 1866 года. Вероятно, Де Фриз сразу понял, что он в своих опытах с кустами ослинника лишь повторил опыты с горохом уже давно покойного к тому времени августинского монаха Менделя и пришел к тем же выводам.
О своем открытии Де Фриз сообщил в номере Comptes rendus de l’Academie des Sciences («Доклады Французской академии наук») от 26 марта 1900 года в короткой заметке «Sur la loi de disjonction des hybrides» («О законе расщепления гибридов»). Едва она была опубликована, как еще два ботаника — Карл Корренс из Германии и Эрих фон Чермак из Австрии — заявили о похожих результатах и успели их в том же году опубликовать. Они тоже, как выяснилось, читали старую статью Менделя и тоже опытным путем пришли к выводу о дискретности наследственных факторов.
Но Де Фриз на тот момент пошел дальше них. Он писал: «И чтобы отдать себе отчет во всех явлениях, надо для каждой наследственной особенности принять отдельную частицу — панген… Эти пангены невидимо малы, однако они совсем другого порядка, чем химические молекулы и их бесчисленные соединения; они должны расти, размножаться и распределяться по всем или почти всем клеткам организма при делении клеток. Они неактивны (латентны) или активны, но размножаться могут в обоих состояниях. Будучи преимущественно латентны в клетках зародышевого пути, они развивают обычно высокую активность в соматических клетках. И именно так, что у высших организмов не все пангены в одной клетке достигают активности, но в каждой одна или же несколько небольших групп пангенов достигают господства и придают клетке ее характер». По образному выражению историка биологии Юрия Чайковского, в этом отрывке вся программа дальнейшего развития генетики.
Де Фриз допустил спонтанную, ничем не мотивированную изменчивость «наследственных корпускул», как их называл Мендель, или «пангенов», как их называл
сам Де Фриз. И пазл строгого научного объяснения эволюции по Дарвину наконец сложился: исходным материалом для естественного отбора служат резкие качественные скачки изменений в наследственных «невидимо малых» частицах — их «мутации», как их назвал Де Фриз.
Родилась теория мутаций, а вместе с ней новая наука — генетика. Про так называемый кошмар Дженкина, который почти полвека держал в подвешенном состоянии дарвиновскую теорию эволюции, можно было забыть. Правда, со временем сами мутации превратились в кошмар для науки, что лишний раз подтверждает нынешняя пандемия коронавируса.
«Кошмар Дженкина»
Как известно, выход в свет в 1859 году фундаментального труда Чарльза Дарвина «Происхождение видов путем естественного отбора, или Сохранение благоприятствуемых пород в борьбе за жизнь» вызвал фурор не только в научном мире. Здесь биология вышла далеко за свои научные рамки, вызвав потрясение устоев общества и почти моментально породив социал-дарвинизм. Но спустя всего восемь лет дарвинизм, победоносно шагавший по планете во всех своих ипостасях — как узкобиологической, так и социальных,— споткнулся о небольшую публикацию в журнале North British Review под названием «Происхождение видов». Ее автором был инженер Генри Дженкин, а ехидство заголовка его статьи становилось понятным по ее прочтении.
Дженкин писал: допустим, что Дарвин прав и с изменчивостью, и с наследственностью, и с естественным отбором, но что из этого следует? Только дикарь, глядя, как ядро вылетает из пушки, может решить, что в конце концов оно долетит до звезд. И скорость ядра, и размах наследственных вариаций стремительно убывают по мере удаления от исходной точки. Поэтому нет никаких оснований ожидать, что выгодные для организма изменения будут накапливаться. Напротив, любое из них очень скоро исчезнет, и вот почему. С кем скрещиваться носителю этого изменения, если вокруг одни нормальные особи? Словом, если следовать логике Дарвина, шансов передать полезное новшество нет: через несколько поколений оно будет «засосано болотом» обычных организмов.
Следовательно, никакая уникальная вариация не может иметь значения для эволюции. Остается предположить одно из двух: либо новая вариация не должна теряться при скрещиваниях, либо она должна возникнуть сразу у значительного процента особей. Однако, писал Дженкин, обе гипотезы отрицают суть дарвинского учения. Первая противоречит наследственности, какой она выглядит в природе и как ее описывает сам Дарвин; вторая же, если ее допустить, приводит к порочному кругу: чтобы распространиться, новая вариация уже должна быть достаточно распространенной.
Более подробно про «кошмар Дженкина» можно почитать в интересных и доступных для понимания читателя без специального образования работах Юрия Чайковского. А сам Дженкин привел пример столь же наглядный, сколь вопиюще неполиткорректный по нынешним меркам: «Представим себе белого человека,
потерпевшего кораблекрушение на острове, населенном неграми… он выживет и станет среди них королем… заведет очень много жен и детей, в то время как множество его подданных умрут холостяками… Но даже его длинной жизни явно не хватит для того, чтобы кто-то из его потомков в каком-либо поколении стал полностью белым… В первом поколении будет несколько дюжин смышленых молодых мулатов, чей ум будет в среднем превосходить негритянский. Нас не удивит, что трон в течение нескольких поколений будет принадлежать более или менее желтокожему королю; но сможет ли поверить кто-то, что население всего острова постепенно станет белым или пусть даже желтым?.. В нашем случае признак попал в исключительно благоприятные условия, способствующие его сохранению,— способствующие, но все же неспособные закрепить и сохранить его».
Из переписки Дарвина известно, что он читал и перечитывал статью Дженкина много раз и в конце концов написал: «Доводы Дженкина меня убедили». Спустя еще три года, в 1870 году, уже в солидном научном журнале Nature (а не в провинциальном общественно-политическом North British Review) вышла статья Альфреда Беннетта под названием «Теория естественного отбора с математической точки зрения». В ней все, о чем писал Дженкин с точки зрения формальной логики, доказывалось строго математически. А вывод был тот же: доказательства эволюции по Дарвину более чем сомнительны. Дарвинизм пошатнулся, но устоял, хотя «кошмар Дженкина», как его назвали, преследовал дарвинизм вплоть по появления теории мутаций Де Фриза.
Новый дарвинизм
В своем первоначальном виде, образца 1901 года, теория эволюции Де Фриза, допускавшая, что один вид переходит в другой скачком в одну мутацию, была крайне несовершенной с современных позиций. Но верным был в ней главный посыл: исходным материалом для отбора служат качественные скачки в наследственных «невидимо малых» частицах. Отталкиваясь от этого, генетики довольно быстро, в течение четверти века, заложили капитальный фундамент под дарвинизм, который стал называться синтетической теорией эволюции.
Стало ясно, где кроется причина ошибки в рассуждениях Дженкина (и Дарвина, кстати, тоже). А крылась она в дискретности наследственного материала — генов, чего ни биологи того времени, ни тем более инженер Дженкин не могли знать. Признаки, закрепляемые отбором, не «разбавляются» при скрещивании, а либо передаются в полном объеме (нивелирующий эффект скрещивания просто не существует, что доказал в своих опытах Мендель), либо не передаются вовсе. Это определяется при скрещивании. Дальше — больше: оказалось, что ген признака может не подвергаться естественному отбору, находясь в рецессивном состоянии. Но новый признак (мутация) не пропадает бесследно в генофонде популяции, хотя его распространение в ней может быть процессом очень длительным и успех не гарантирован.
Общепринято, что окончательно связал эволюционное учение Дарвина и законы наследственности, установленные к тому времени генетиками, и тем самым покончил с «кошмаром Дженкина» советский генетик Сергей Четвериков в статье «О некоторых моментах эволюционного процесса с точки зрения современной генетики» 1926 года. За
границей эта его статья никогда не публиковалась, но западные генетики ее знали и читали ее переводы с русского благодаря коллегам Четверикова Феодосию Добржанскому и Николаю Тимофееву-Ресовскому. Оба они были «невозвращенцами»: Добржанский работал с 1927 года в США, Тимофеев-Ресовский — с 1925 года в Германии.
По Четверикову, теория эволюции Дарвина теперь выглядела так: мутация (насыщение видов в природе возникающими мутациями) — свободное скрещивание — естественный отбор. При этом он сделал допущение, что мутационный процесс в природе протекает точно так же, как и в условиях лаборатории. Поэтому ученые вправе экстраполировать выводы из результатов лабораторных опытов на природные ситуации.
Первый такой вывод — это непрерывное во времени возникновение новых мутаций у всех видов живых организмов. Второй — рецессивность большинства вновь появляющихся мутаций по отношению к аллелям (формам гена) дикого типа, распространенным в природных популяциях. Третий — в природе преобладает панмиксия (свободное скрещивание). На этих трех китах до сих пор стоит наука «популяционная генетика», и дополнительных опор ей пока не нужно.
Мутации — народному хозяйству!
Сегодня «кошмар Дженкина» не более чем любопытный факт из истории развития наших представлений об эволюции жизни на Земле. А вот мутации вышли далеко за пределы академических исследований эволюции. Микроорганизмы, которые отличаются относительной простотой их организации и стремительной сменой поколений по временным меркам человеческой жизни, оказались идеальными модельными объектами для генетиков, изучающих мутационные процессы.
У микроорганизмов достаточно легко наблюдать и индуцировать самые разнообразные мутации. Гаплоидность (одинарный набор хромосом) многих микроорганизмов обеспечивает проявление рецессивных мутаций, которые у высших диплоидных (с двойным, дублирующим набором хромосом) организмов могут быть замаскированы присутствием нормальной аллели. Микроорганизмы легко культивировать и получать от одной исходной клетки колонию генетически однородных клеток, а затем размножать их до количества, необходимого для биохимического и молекулярного анализа. Отсюда до практического использования полезных, с точки зрения человека, мутаций бактерий был один шаг.
Из более чем 100 тыс. видов известных на сегодня микроорганизмов человеком используются несколько сотен, и число их растет. Они продуцируют десятки веществ — аминокислот, белков, антибиотиков, витаминов, липидов, нуклеиновых кислот, ферментов, пигментов, сахаров и т. п. На свойстве живого мутировать выросла целая отрасль — микробиологическая промышленность.
Оборотной стороной мутагенности микроорганизмов стало появление устойчивых к антибиотикам бактерий — супербактерий (superbugs). Не каких-то новых бактерий, а самых обычных — например, кишечной палочки. По оценке Всемирной организации
здравоохранения (ВОЗ), сейчас смертность от таких бактерий-мутантов колеблется от 700 тыс. до нескольких миллионов в год. Иначе чем кошмаром для медицины это трудно назвать.
Кошмарная генетика вирусов
Еще проще устроены фаги и вирусы. Вирусы имеют очень небольшой период размножения и повышенную частоту мутаций: одна или больше мутаций на геном за один раунд репликации РНК вируса. Образно говоря, постоянное мутирование и есть образ их жизни. В этом году даже появилась теория врожденной малограмотности вирусов: они, мол, в принципе не могут без ошибок скопировать при размножении собственный геном.
Но теории теориями, а в реальной жизни повышенная мутагенность вирусов не дает возможности раз и навсегда покончить даже с банальным гриппом. Пока после очередной сезонной эпидемии гриппа готовится очередная вакцина против него (обычно это занимает не менее полугода), вирусы гриппа успевают мутировать, и, как говорится, на колу мочало, начинай сначала. Это самый страшный секрет Полишинеля ежегодных кампаний вакцинации граждан от гриппа: нас вакцинируют всегда против прошлогоднего гриппа (в лучшем случае — прошлогоднего, а то ведь на складах остается много вакцин и за предшествующие годы, на всякий случай). А эффективность вакцинации зависит от того, насколько вирус гриппа этого года похож на вирус прошлогоднего гриппа. Когда уж очень непохож, возникают пандемии «свиного», «куриного», «гонконгского» и прочих сильно мутантных гриппов.
Против других ОРВИ, которые протекают в легкой или средней тяжести формах, ученые даже не стараются создать вакцины. Почихал, покашлял человек недельку — и слава Богу, теперь у него есть собственный иммунитет против данного вируса. Когда зимой 2002–2003 годов появился ТОРС-коронавирус (SARS), вызвавший эпидемию атипичной пневмонии со смертностью 10%, а в 2012 году — БВРС-коронавирус (MERS) со смертностью 34%, ученые было начали готовить против них вакцины, но едва лог-фазы эпидемий закончились, бросили это дело. Они понимали, что в следующий раз будет что-то другое. Правильно понимали. Теперь тоже вроде бы SARS-коронавирус, но уже с другими мутациями — SARS-CoV-2. Вакцина 2003 года, если бы ее тогда не бросили на полпути, а доделали бы до конца, едва ли спасла бы от него.
Что касается многоклеточных организмов, то ученые уже сравнительно давно начали пересаживать полезные человеку с практической точки зрения гены от одного вида к другому, или, проще говоря, создавать полезные мутации. ГМО сейчас обычный компонент нашего с вами рациона. А с недавних пор ученые даже научились удалять хирургическим путем (методами молекулярной хирургии) врожденные вредные мутации из генома человека. Здесь у ученых все в порядке. Теперь их опять «кошмарит» механизм эволюции. На этот раз — эволюции вирусов, где в теории все гладко, а на практике — хуже некуда. Здесь своего Хуго Де Фриза пока не нашлось.